四、不是“残酷竞争”而是“共生互惠”
4.1 竞争思想的由来
达尔文进化假说的支柱之一,来源于英国经济学家和人口学家托马斯‧马尔萨斯 ¹⁴²(Thomas Malthus, 1766—1834)在1798年提出的“生物过度繁殖论 ”¹⁴³,认为生命过度繁殖、人口过度增长,所以才需要竞争、才需要自然选择和淘汰。
马尔萨斯(Thomas Robert Malthus)在《论人口原理,因为它影响社会的未来进步》(An Essay on the Principle of Population, as it Affects the Future Improvement of Society)一书中提出,呈代数级数增加的食物供给,赶不上以几何级数增长的人口,供需不平衡将无可避免地使部分人遭受被淘汰的悲惨命运。
在达尔文的年代,马尔萨斯的理论已跻身英国主流思想。1838年,29岁的达尔文正在思考演化可能的机制。达尔文将马尔萨斯的“生存竞争”引入“进化论”,他在《物种起源》中表示,生命不论同种或异种,都面对着生存竞争,因此生命繁殖得多,能活下来的却没那么多。自然界生物之间为了生存而争夺有限的生存资源,包括生存空间、食物、配偶等等,作为强势个体或群体得以生存的基础。
4.2 生物间“共生互惠”挑战“竞争法则”
大自然中,某些生物个体之间互相竞争食物的现象的确存在,可是,达尔文错在把它绝对化、极端化地描述成为一种规律,用这种弱肉强食、残酷竞争的冷酷关系定义自然生物之间的关系。但自然界生物之间的相互依存、相互支持、共生互惠 的这种更普遍存在的更加本质上的关系¹⁴⁴,却被达尔文有意忽略掉了,而后者其实早就被科学家观察发现到了。
早在1877年,德国植物生理学家弗兰克(Albert Bernhard Frank)使用曾经用于描写共同生活在一个社区的民众的词语“共生 ”(symbiosis),来描述地衣中真菌与藻类的互惠共生关系。1879年德国植物学家德巴利(Henrich Anton de Bary)概述了共生的三个标准 :两个不同物种;它们生活在一起;它们紧密接触。¹⁴⁵
20世纪中期,生态学研究有了飞速发展,生态系统 ¹⁴⁶的研究成果告诉我们:自然界的任何物种或个体都处于一定的生态系统中,生态系统内的物种与物种之间、生物与环境之间的互相依存、互利互惠关系,是自然界的普遍规律 ¹⁴⁷。
美国国家科学院院士兰斯基在他的大肠杆菌实验中曾经观察 到这样的现象,同一个烧瓶中出现了两种类型的大肠杆菌,一种繁殖能力相对较弱,另一种相对较强。按照达尔文的自然选择、优胜劣汰的理论,繁殖能力较强的细菌会最终代替繁殖能力较弱的细菌。但令兰斯基惊讶的是,实验结果是两种类型的细菌相安无事、共同生存,弱者并没有被淘汰。¹⁴⁸
达尔文的进化假说相当于足球的淘汰赛,最后胜出的往往是最强大的;实际上,生物之间的关系不是这样的竞赛关系,而是讲究“生物多样性”的,每一个生物个体都有其独特特点,大多数物种都能共同繁荣发展。
第二,科学研究还发现,生活在同一地区的植物种类越多,植物总体上的生长趋势越兴旺 ¹⁴⁹。
数百项关于陆地、水生和海洋生态系统的研究表明,具有更多生物种类的生态系统,即具有更多生物多样性 的生态系统的生物生产率(productivity)──定义为“生物量(biomass)的产生率”(通常以单位时间内每体积的质量单位表示,例如每天每平方米克数)──约是栽培单一物种的生态系统的两倍,而且这种差异会随着时间的推移而加大¹⁵⁰。生态系统中的生物多样性带来的益处,与物种之间互补、更有效地利用有限资源、增加抵抗疾病的能力等等多种原因有关。
这些生物多样性的理论和实验都具有共同特点,都是允许多种不同的竞争物种长期共存 。
因此,种种关于共生现象的科学研究,有力地挑战 了达尔文关于“自然竞争、适者生存”的主要核心假设¹⁵¹。
第三,地球上的生物变化万千、奇特多姿,却彼此配搭得十分巧妙。绿色植物的光合作用把太阳能变为植物体内的化学能,直接或间接地为所有的动物提供了食物;而动物的排泄物和遗体又是植物生长的原料。在生物酶的催化下,植物在常温、常压下可合成各种复杂的有机物。一切生物呼吸时,需要吸收O₂,吐出CO₂,植物的光合作用则用CO₂合成各种有机物,同时释放出O₂。这些巧妙安排,难道仅仅是凑巧的结果吗?
一个生物的复杂高效系统尚且不可能随机变异而来,而由彼此之间存在着巧妙合作的、如此众多的生物共同组成的生态系统,又怎么可能是随机突变的结果呢?
第四,共生是生态群落建立和维持的主要机制。如果破坏生物之间自然存在的、共生互惠的关系,破坏生态环境,可能会导致生物界的灾难。
举个例子,用传统的种植方式种蔬菜水果,既不需要化肥,也不需要农药。因为自然界之中也有生态链,虫子有虫子的天敌,比如青蛙、小鸟。如果人为介入,用农药消灭虫子,青蛙、小鸟就没有食物了,相当于破坏了大自然的生态平衡。
所以,杀虫剂 破坏了生物之间的共生互惠的关系。杀虫剂除了杀死昆虫或杂草外,还可能毒害许多其它生物体,包括鸟类、鱼类、益虫和非目标植物;农药如果破坏土壤周围的生态链,最终的结果也会给人带来伤害。¹⁵²
随着科学技术的发展对生态环境的破坏和污染,人们越来越意识到要吃有机食品 。所谓“有机”,就是在农业种植或动物养殖过程中,不允许使用人工合成肥料、大多数合成杀虫剂、抗生素或生长激素,并禁止用辐射、基因工程技术改变作物的基因,禁止使用化学防腐剂等合成物质。这样生产和加工出的农副产品和加工食品,才能算作有机食品。¹⁵³
所以,不管从哪个逻辑上去分析,共生现象都不符合达尔文进化假说的核心思想。生物物种的共生互惠是对进化假说真正的挑战。
4.3 人与微生物间“共生互惠”
物种之间存在生态平衡的关系,植物动物之间存在着相互关系,人自出生之日起就与微生物建立了相互之间的紧密关系。
人体并非是一个单纯的个体,而是由体内的微生物细胞和人体细胞共同构成的“超级生物体”。人体是一个复杂的微生态系统,我们的身体里居住着数以万亿计的微生物 ,包括古菌(又称古细菌、古生菌)、细菌及病毒,分布在我们的皮肤、口腔、鼻腔、生殖器及肠道等部位,构成了被称为人类的第二基因组的人类微生物组。¹⁵⁴
其中,胃肠道的微生物细胞总数约10¹⁴个 。人类与肠道细菌建立了互惠共生体系:前者为后者提供了适宜的温度、丰富的营养物质、优越的栖息和繁殖生境(物种群体赖以生存的生态环境);后者则能够调控肠道上皮细胞发育、防止细胞损伤、调节宿主脂肪存储、刺激肠道血管生成、构建生物膜等免疫屏障、阻抑外来致病菌的侵入定殖、帮助人类消化大分子食物成分、参与糖和蛋白质代谢、合成人类必需的维生素与氨基酸等,并促进营养元素被人体吸收利用。
现代科学研究证明 ,肠道微生物与人大脑之间形成了微生物—肠—脑轴(Microbiota-gut-brain Axis)体系,与人体各种疾病(包括免疫、癌症、心血管、神经退行性疾病等等)均密切相关,甚至与精神活动也有密切关系。这些精神活动包括社会行为、压力反应、认知等方面。与我们人类共生的这些微生物,深刻影响和调控着人体,为人类健康做出了无可替代的重要贡献。
在医学治疗中,如果过度使用抗生素,容易“滥杀无辜”,导致人肠道菌群失调。这对人体的免疫力和大脑都会带来负面影响,甚至引发疾病。
高盐饮食也会抑制肠道内的正常菌群生存,导致人肠道菌群失调,产生免疫功能紊乱 等各种疾病¹⁵⁵。
在当今地球上,细菌、植物、动物、人类之间,均依靠其共生体系及多种体系共存、发展和繁荣。在构成生态系统的同时,生物多样性也得以增加、保护和维持。
总之,生物共生是自然界生物群体包括人类在内维持相互关系的一个普遍法则。所以,达尔文的进化假说强调生物物种之间通过竞争而进化,从根本上是错误的。竞争绝不是生命的主旋律。
4.4 基因并非“自私”而是“利他”
进化假说的提出者带着自私的理念来看待这个世界,将生物基因属性描绘成自私的,认为一切都是自私的基因导致的结果。
英国进化生物学家和作家理查德‧道金斯(Richard Dawkins, 1941—)在《自私的基因》 (The Selfish Gene)一书中指出¹⁵⁶,“基因是自私的基本单位。基因具有潜在的永生性,使其成为自然选择基本单位的良好候选者。”“我认为,成功基因的主要特质是无情的自私。这种基因的自私通常会导致个体行为的自私。”“经过几代的自然选择,‘利他群体’会被自私群体超过,并且难以与自私群体区分。”
他们甚至还认为,一只鸟或一只蜜蜂冒着生命和健康的危险将其后代带到这个世界上,不是为了帮助自己,也不是为了帮助它的物种,而是为了让它的基因继续存在 ¹⁵⁷。
道金斯实际上是把达尔文提出的生物个体之间的竞争归结到不同版本的基因(称为“等位基因”)之间的竞争关系。他的观点是把达尔文的进化论往基因的微观层面推进一步,但是事实情况真的是这样吗?
道金斯提出这一理论之后,遭到了科学家的质疑。
1999年《复杂系统进展》(Advances In Complex Systems)期刊中,美国复杂系统数学分析应用专家、新英格兰复杂系统研究所所长亚尼尔‧巴-亚姆(Yaneer Bar-Yam, 1959—)博士认为,自私基因理论存在致命缺陷 ¹⁵⁸,使用自私基因模型研究 的生物现象,存在重大偏差,只能应用于有性生殖导致等位基因完全随机交配的群体。这样的群体在生物学中被称为“随机交配”(panmictic)种群,俗称“盲婚哑嫁”。但其实许多生物种群并不满足这一条件。因此,道金斯把生物群体过分简化了,不符合现实,他所提出的以基因为中心的观点并不能代表生物种群的真实状态。“自私基因”的概念不成立。¹⁵⁹
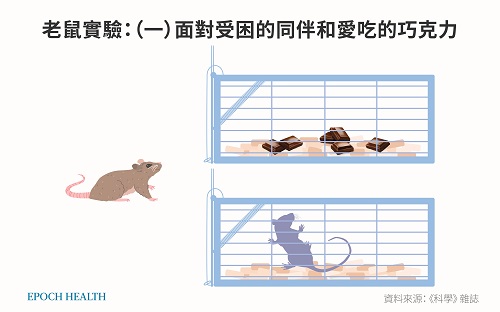
2011年《科学》(Science)杂志发表了一项有趣的动物对照研究 ,实验人员将一只大鼠A关进铁笼子里面,观察外面一只自由活动的大鼠B的行为。这个实验重复测试了30只大鼠B在上述场景中的行为,结果发现,自由活动的大鼠B围绕着笼子的活动频率明显增加,它们想尽一切办法打开笼子,直到营救成功。它们的这种营救行为与在没有被关大鼠A的空笼子外面时的对照场景相比,具有显着的统计学意义。¹⁶⁰
另外,如果没有其它特殊情况发生,大鼠可以吃掉至少7块巧克力,而不去吃其它普通的食品。
实验人员把自由大鼠B放到两个笼子前面,其中一个笼子里,另一只大鼠A被关在里面;在第二个笼内放着大鼠爱吃的巧克力。一边是美食的诱惑,一边是受难的同伴,猜猜看这只大鼠B会做什么?
实验人员看到了更令人惊叹的一幕,老鼠B并不自己独享巧克力,相反,它会以几乎相同的速度打开两个笼子(因为打开两个笼子需要时间、学习和实践),之后还会拿出自己专门攒下的30%的巧克力分给“受难的同伴”。
大鼠面对受难的同伴和美食的诱惑,做出无私的选择。 (大纪元制图) 按照道金斯的理论,他把基因和个体的行为等同起来,认为基因就是老鼠、老鼠就是基因。可是现在这些重复对照的科学实验结果显示,老鼠在利益面前表现出来的是利他的行为。如果老鼠的行为不自私,那么老鼠的基因的主要特质也不可能是自私的。
黑猩猩之间会相互梳理毛发,并与其它伙伴分享食物。相互梳理毛发可以为双方带来直接利益,而分享食物通常是为了联络感情。不过,黑猩猩的这些合作行为,人们不清楚在多大程度上是为了让对方获利还是最终为了谋求自己的私利。
2017年《美国国家科学院院刊》(Proceedings of the National Academy of Sciences, PNAS)发表了一项研究 ,题目是“黑猩猩以牺牲自身个体的利益回报恩惠”(Chimpanzees return favors at a personal cost),用黑猩猩(Pan troglodytes)做了多次重复的实验,给我们带来很多启发。¹⁶¹
“伴侣黑猩猩”首先接受一项培训,学会放弃一个自己独占食物而导致“受试黑猩猩”空手而归的选项,同时冒着失去食物的风险,把选择食物的权力交给“受试黑猩猩”。也就是说,“伴侣黑猩猩”首先接受了利他(altruism)行为的训练。
“受试黑猩猩”当看到“伴侣黑猩猩”冒着失去食物的风险也要帮助自己时,为了回报对方的恩惠,在伴侣利他行为的带动下,也更容易做出牺牲自己的物质代价而让伴侣受益的行动。这个结果具有显着的统计学意义。
在不同场景对比的实验中,研究者发现,“受试黑猩猩”可以清楚地理解和区分“伴侣黑猩猩”的行为动机,能将真正的亲社会行为(prosocial)也就是真正的“利他”行为与潜在的自私行为区分开来。
从这个实验中我们看到,即使是动物,也有学会为了帮助同伴而放弃自己利益做出无私行为的能力,并且一个个体的利他行为能正面影响其它个体也做出利他的举动。那么按照道金斯的理论,如果基因都是自私的,这些动物如何能做到为了帮助同伴而牺牲自己的利益?这显然不是道金斯的逻辑所能涵盖的。
其实,不仅基因不是自私的,而且动物的行为也往往不是自私的,而是利他的。根据常识,利他现象 在社会性昆虫、鸟类、哺乳类乃至人类社会中尤为突出¹⁶²。所谓“自私的基因”完全是没有根据的说法。

 您的位置:
您的位置: